 Lignende trekk
Lignende trekk
(Fra boka 'The Design of Life' kap 5.1-4; av W. Dembski og J. Wells)
1.Klassifikasjon og fortolkning
De fleste lignende trekk i forbindelse med organismer, kan deles i arrangementer av grupper innen grupper. Desto flere likheter som deles, og desto større grad av likhet, desto nærmer klassifiserer vi organismer som deler dem:
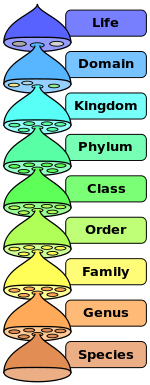
En hund er mer lik arten ulv, enn arten rev, derav følger at hunden og ulven klassifiseres i samme slekt (genus: Canis), og reven i en annen slekt. Likevel er en hund mer lik en rev, enn en katt, så de to første klassifiseres i samme familie (family: Canidae), og katten i en annen familie. Men en hund er mer lik en katt enn en hest, så de to første er plassert i samme orden (Order: Carnivora), og hesten plasseres i en annen. Likevel er en hund mer lik en hest enn en fisk, så de to førstnevnte plasseres i samme klasse (class: Mammalia), og fisken i en annen klasse. Likevel er en hund mer lik en fisk enn f.eks. en orm, og de to førstnevnte plasseres i samme rekke (phylum: Chordata), og ormen tilhører en helt annen rekke. Likevel har hunden mer likhet med en fisk, enn f.eks. et eiketre, derfor kategoriseres hunden og fisken i samme rike (kingdom: Animalia), og treet i et annet rike (planteriket).
For Darwin var likhet et resultat av felles avstamning. Han fortolket lignende trekk som 'familie-likhet': to organismer er like fordi de nedkommer fra en felles stamfar. Neo-Darwinister (ND) hevder grader av familielikhet avslører hvor nært organismene er beslektet til en felles stamfar. F.eks. er alle pattedyr bygd på en felles kroppsplan. ND fortolker dette slik at alle pattedyr nedstammer fra en felles stamfar, i klassen pattedyr, som hadde den kroppsplanen. Ulikheter mellom pattedyr sies således å avsløre hvordan basis-planen har blitt tilpasset i hver art, under press fra naturlig seleksjon. For ND er likheter generelt en konsekvens av slektskap.
Bilde 1. Oversikt over taksonomiske klasser
Fordi fossiler ikke er levende organismer, kan vi ikke direkte benytte dem til å etablere avstamnings-relasjoner og avstamning. Forestill deg at en graver opp to menneskelige skjelett av nyere dato. Uten å identifisere merker og skriftlige nedskrivninger, kan vi ikke fortelle hvordan de to er i slekt med hverandre. (Eneste unntak ville være om vi ekstraherte identisk DNA fra dem, i det tilfellet ville de være eneggede tvillinger). Om vi ikke kan si hvordan to skjelett av nyere dato er beslektet, kan vi heller ikke fortelle hvordan gamle fossiler av ulike arter er relatert, eller om de er det i det hele tatt.
Som en konsekvens må paleontologer stole på likheter for å konstruere hypoteser om evolusjonært slektskap. I følge ND teori: desto større antall likheter mellom de to organismene, desto nærmere evolusjonært slektskap er det sannsynligvis. Men å avsløre og fortolke likheter er ikke så enkelt som det kan høres. Så snart vi fortsetter videre enn heller åpenbare likheter, som at fugler har fjær og fisker har skjell. Det er ikke alltid enkelt å avgjøre hvilke organismer som burde klassifiseres sammen. Likheter opptrer i et lappeteppe-mønster,  som gjør klassifikasjon vanskelig.
som gjør klassifikasjon vanskelig.
Bilde 2. Kenguru -eks. på pungdyr
Betrakt pungdyrene, som fullfører embryo-utviklingen i en utvendig pung på morens mave, i motsetning til pattedyr med morkake (placenta), slik mennesket har. Dyr med morkake og pungdyr, er noen ganger slående like. F.eks. er nordisk ulv og den nå utdødde tasmanske ulven, veldig like, f.eks. i skjelettstruktur. Det samme gjelder levesett og oppførsel. Likevel skilte de to dyrene seg fundamentalt i sin tidlige utvikling. Til tross for den slående likheten mellom voksne, fokuserer taksonomer (som klassifiserer dyr) på forskjellene, og klassifiserer dem i ulike kategorier. Dermed klassifiserer de ulven i lag med hund i samme slekt (genus: Canis), mens den tasmanske ulven klassifiseres i lag med pungdyr som kenguru. ND fortolker denne anatomiske forskjellen til å indikere at de to typene ulver bare var fjernt beslektet, og at hver hadde en lang og atskilt evolusjonær historie, som går tilbake til da Australia ble et eget kontinent.
Forutsetningen fra ND er at de selektive regimene og omgivelses-nisjene som produserte den nordiske ulven, var i nærheten like av det i Australia. Dermed, ved å tilpasse seg lignende omgivelser kom de to helt atskilte ulvene til å ligne mer og mer, inntil de ble tilnærmet nesten identiske. Likevel er det problemer med denne måten å resonere på: i) bevisene støtter ikke formodningen at de omgivelsene for begge ulver var lignende og ii) Det er ingen grunn til å tro at selv med lignende omgivelser, så ville to separat utviklende arter utvikle ikke bare ett lignende trekk, men en hel pakke av lignende trekk, som matcher punkt for punkt. Men sammentreffene stopper ikke der. Australia er hjem for en skare av andre pungdyr, som har sine motparter i pattedyr: pungdyr: katter, ekorn, villsvin, maurslukere, moldvarper og mus. Dermed blir det to hele underklasser av pung- og patte-dyr på to ulike kontinenter, som utvikler de samme morfologiske typene, og begge deler hele pakker av lignende trekk. Uten noen form for teleologisk styring, krever konvergent evolusjon en opphopning av sammentreff på sammentreff, som tyner troverdigheten.
 2. Analogi og homologi
2. Analogi og homologi
Pungdyr reiser et interessant spørsmål for taksonomer: Om likhet er basis for klassifikasjon, hva skulle vi gjøre dersom ett sett likheter klasjer med et annet sett? Selv om den tasmanske ulven var slående lik den nordiske, i de fleste egenskaper og utseende, så er den et pungdyr og herved lik kenguruen i denne vesentlige egenskapen. På hvilke lignende trekk, skal vi dermed bygge vårt klassifikasjonsskjema?
I biologi kommer lignende trekk ofte i to hovedformer: funksjonelle og strukturelle. F.eks. blir fuglevinger og insekt-vinger benyttet til flyvning. Begge fungerer på samme måte: luftstrømmer løfter nedenfra mot vinge-overflaten for å forsyne et løft, og å slå med vingene gir skyv. Likevel er den interne strukturen til en fugle-vinge helt annerledes enn i en insekt-vinge. Fuglevingen består av kjøtt, forsynt med næring og oksygen via et nettverk av blodårer. Dens strukturelle stabilitet kommer fra ben på innsiden. Innsektvingen på sin side, har verken ben eller blodårer. Den består av en tynn membran tett spunnet rundt ett nettverk av trådstrukturer, i likhet med en drage. Fuglevinger og insektvinger tjener samme funksjon, men er meget ulike i struktur. Her er likheten funksjonell heller enn strukturell. Men det er ikke eneste mulighet. Likhet i struktur og ulikhet i funksjon er vanlig. F.eks. ben-mønsteret i en flaggermus sin vinge, er like det i en delfins finner, selv om vingen benyttes til å fly med og finnen er for svømming.
Hvilken likhet er mest relevant for biologisk klassifisering -likhet i funksjon eller likhet i struktur? C. Linné møtte dette problemet på slutten av 1700-tallet, og valgte å klassifisere organismer i henhold til strukturell i stedet for funksjonell likhet. Således klassifiserte han flyvende insekter i lag med andre insekter, på grunn av deres strukturlikhet, og ikke sammen med fugler. PÅ 1840-tallet kalte den britiske anatomen R. Owen funksjonell likhet 'analogi', og strukturell likhet 'homologi'. Owen mente distinksjonen som en hjelp i forhold til Linné's klassifikasjonsskjema: analogi indikerer uavhengig tilpasning til eksterne forhold, mens homologi indikerer strukturell beslektethet. Både Linné og Owen så strukturelle likheter som mer tillitverdig guide for å klassifisere organismer, enn funksjonelle likheter.
Bilde 3. Eks. på 'analoge' strukturer
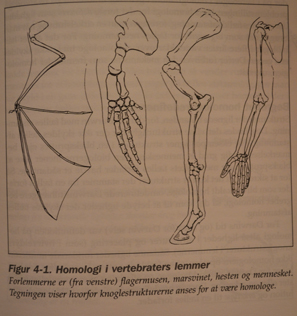
Det klassiske eks. på homologe strukturer, er forbena til virveldyr. Selv om en flaggermus har vinger til å fly, og en tannhval har finner for å svømme, en hest har ben til å løpe, og et menneske har hender til å gripe, så er benmønsteret i deres forlemmer lignende. Slike skjelett- messige likheter sammen med andre slektskap som varmblodighet og melkeproduksjon, rettferdiggjør å klassifisere alle disse som pattedyr, til tross for sine eksterne ulikheter. Lik andre pre-darwinistiske biologer, betraktet Owen homologe trekk til å bli oppnådd fra et felles mønster, eller 'arketype'. En slik arketype kunne forstås på ulike vis, f.eks. som en 'Aristotelisk form', iboende i naturens struktur, eller en prototype organisme, blant annet. Owen og Darwin så arketypen som en 'prototype' organisme. Owen så organismene som konstruert ut fra en felles plan. Darwin så dem som nedstammet fra en felles stamfar.
I følge ND forsyner homologier bevis for evolusjonær avstamning. Således fortolker ND likhet i skjelettstruktur, som bevis for at de har nedstammet fra en felles sti. Ta menneskehanda og hundens forben. Begge inneholder de samme typer og bein, og bena til begge er tilordnet de samme navnene. Selv om benstrukturen er lik, er de individuelle bena ganske ulike i form og funksjon. Hunden kan ikke gripe objekter med tærne, eller holde tommelen mot resten av tærne. Menneskelige hender er ikke laget for å gå eller stå på i lengre tid. Deres likhet til hundens poter, er dermed uavhengig av funksjon. I følge ND er eneste alternativ at deres likhet skyldes arv fra en felles stamfar, som besatt denne basis-arrangering av ben.
Hva var denne felles stamfaren? I følge Darwin var det en form for Insectivore- ordenen av pattedyr som inkluderer moldvarper og spissmus. Dette dyret er så antatt å ha utviklet seg fra et opprinnelig krypdyr, som i sin tur mottok sine forlemmer fra en semi-akvatisk fisk, som formodentlig benyttet sine ben-finner til å dra seg fra sølepytt til sølepytt under sesongpreget tørke. En finne synes ikke som det beste materialet for en  hests ben eller for en hunds pote, men det var eneste tilgjengelig råmateriale (og evolusjon kan bare jobbe med tilgjengelige materialer). Følgelig er både et menneskes hand og hunds pote homologe til en fisks finne. I det den mangler nye ideelle deler, må ND-mekanismene forme og reforme eksisterende deler.
hests ben eller for en hunds pote, men det var eneste tilgjengelig råmateriale (og evolusjon kan bare jobbe med tilgjengelige materialer). Følgelig er både et menneskes hand og hunds pote homologe til en fisks finne. I det den mangler nye ideelle deler, må ND-mekanismene forme og reforme eksisterende deler.
Bilde 4. Mulig stamfar i følge Darwin
Mens ND har tilbudt mange rasjonaliseringer for hvorfor gap skulle eksistere i fossil-registeret, så fortsetter de å lete etter mellomliggende fossiler, som bevis for sin teori. Likevel er ubrutte serier av fossiler som viser nedstamning av en art fra en annen, aldri funnet. Siden det er slik, blir homologi den eneste måte en kan realisere evolusjonært slektskap på. Men dette innebærer at ND må fortolke analogier (funksjonelle likheter), som ikke også er homologe (strukturelle likheter), som resultat av konvergent evolusjon; formodentlig som tilsvar på miljømessige likheter. Følgelig inntraff evolusjon av flyvge-evne hos insekter og fugler, ved uavhengige prosesser som konvergerte som tilsvar på lignende omgivelses-press.
Likevel er ikke distinksjonen mellom homologi og analogi lette å trekke. Homologe strukturer kan være helt ulike i både form og funksjon (eks. en flaggermus' vinge og en hests frambein).På den annen side kan meget likt utseende strukturer som utøver samme funksjon, kun være analoge og således irrelevante for klassifikasjon og dragning av evolusjonære slektskap. Linné klassifiserte hval som fisk, i det han 'ikke innså at deres fiske-lignende form, ikke var en homolog men bare en analog likhet' (kommenteres under). Gjentatte ganger i studiet av taksonomi, er strukturer med forbløffende likhet først betraktet som homologe, men ble senere betraktet som bare å være analoge. Mange strukturer er faktisk blandede, og er både homologe og analoge.
 3. De forbløffende pandaene
3. De forbløffende pandaene
Kjempepandaen og den mindre røde pandaen, illustrerer problemet med å skjelne homologe og analoge strukturer. Begge pandaer bor i bambus-skogen i Sør-vest Kina. I over ett århundre, har forskere som har studert de to pandaene, ikke vært enige i hvorvidt de er med i familien bjørner eller vaske-bjørner (racoons). Siden de første forsøk i 1869 er det publisert over 40 større vitenskapelige studier om emnet. Det forbløffende er at disse studiene har splittet det vitenskapelige miljøet omtrent på midten. En vitenskapsmann beskrev denne mangelen på klassifikasjon som et taksonomisk sett ping-pong. Så ble saken løst til de fleste biologers tilfredshet, ved at kjempepandaen ble klassifisert som en bjørn, og likhet til den er blitt bekreftet av senere studier. Samt at den røde pandaen, er klassifisert som en vaskebjørn. [Dw. Davis, 1964). Konsekvensen av rådende syn blant biologer, blir imidlertid at pandaens tommel utviklet seg uavhengig av hverandre, to ganger ('konvergent evolusjon').
Men å ha en eneste vaskebjørn utenfor vestlig halvkule, slo mange biologer som usannsynlig. Det ville være mer sannsynlig om begge var bjørner, eller begge var vaskebjørner, mente de. Med til historien hører at kjempepandaen har 42 kromosomer, mye mer i likhet med den røde pandaens 36, enn til de 74 kromosomer i de fleste bjørner. Kjempe-pandaens 'tommel' gir en fingerferdighet, som ikke er funnet blant andre bjørner. Denne strukturen fungerer som en imøte-gående tommel, selv om den ikke er en skikkelig tommel og bare delvis kan 'mot-stilles' de andre fingrene. Dens evne til å håndtere og strippe bambus, er bemerkelsesverdig og forbruker mye av dens tid, og dens 'tommel' er nøkkelen til suksess i denne aktiviteten. Denne strukturen samt dens støttestruktur, er også felles i begge pandatypene, selv om den røde ikke har den i samme grad som kjempepandaen. I tillegg har begge pandatyper flere like trekk av oppførsel. F.eks. er det ingen av dem som går i dvale.
Sammentreff som disse, og det er mange i den virkelige verden, reiser spørsmålet hvorvidt lignende trekk noensinne gir tillitverdig informasjon om evolusjonært slektskap.
Bilde 5. Pandaens 'tommel'
 4. ND redefinisjon av homologi
4. ND redefinisjon av homologi
I 'Origins of Species' argumenterte Darwin at den beste forklaring på homologi, er nedstamning med modifikasjoner. Darwin så homologi som et viktig bevis for sin teori, og listet det blant "de faktorer som 'proklamerer så tydelig at de utallige arter, slekter og familier som jorden er befolket av, alle nedstammer innenfor sin egen klasse eller gruppe fra felles foreldre." (3) Men noen trekk er ikke oppnådd gjennom felles avstamning, som vi så i tilfellet med pungdyr og placenta-pattedyr. Vi har også nettopp sett at biologer ikke lenger mener at pandaens tommel kan forklares ved felles slektskap i forholdet mellom kjempe- og den røde- pandaen. Likedan er strukturen i en blekkspruts øyne mye lik strukturen i ett menneskes øye, men biologer tror ikke likheten skyldes felles avstamning. De ser nå slike lignende strukturer som resultat av 'konvergent evolusjon'.
For å sikre at bare strukturer som skyldtes en felles stamfar, skulle kalles homologe, redefinerte dermed ND homologi til å bety likhet som skyldes felles avstamning. I følge Ernst Mayr som er sjefsarkitekten til ND, så er det "etter 1859 bare en definisjon av homologi som gir biologisk mening.. Å tilordne to organismer som homologe når de er oppnådd gjennom et lignende karakteristika fra en felles stamfar.' (4) Selv etter at homologi ble redefinert, var imidlertid ND ufullstendig uten en mekanisme til å forklare hvorfor homologe trekk var så like i så ulike organismer. Da ND oppsto i 1930-40 åra, lovte den å løse dette problemet: 'homologe trekk var resultatet av lignende gener arvet fra en felles stamfar. Men biologer har visst i flere tiår at homologe trekk kan oppstå fra ulike gener, og at lignende gener kan ligge under ikke-homologe trekk. Så mekanismen som produserer homologi er ukjent (mer på CD-en som følger boka).
Bilde 6. Forklarer det som skal defineres
Videre hvis homologi defineres som likhet som skyldes felles avstamning, så kan ikke homologi benyttes som bevis for felles avstamning (sirkelargumentasjon). En tilhenger av ND som vil avgjøre hvorvidt virveldyrs fram-lemmer er homologe, må først bestemme hvorvidt de nedstammer fra en felles stamfar. Det må altså være bevis for en felles stamfar, før lemmer kalles homologe. Men så å snu argumentet rundt, og argumentere for at homologe lemmer indikerer felles avstamning, skaper en ond sirkel: Felles avstamning etablerer homologi, som igjen etablerer felles avstamning.
 Flere biologer og filosofer har bemerket og kritisert denne sirkellogikken. I 1945 skrev J.H. Woodger at den nye definisjonen 'satte vogna framfor hesten." (5) A. Boyden påpekte i 1947 at ND homologi først krevde at vi fant opphavet og så besluttet at de korresponderende organer eller deler" er homologe. "Som om vi kunne vite opphavet uten de essensielle likhetene til å lede oss!" (6) Da ND-paleontolog G.G. Simpson forsøkte å benytte homologi som begrunnelse for felles stamfar, beskyldte biologer R. Sokal og P. Sneath ham for 'sirkulært resonnement.' (7)
Flere biologer og filosofer har bemerket og kritisert denne sirkellogikken. I 1945 skrev J.H. Woodger at den nye definisjonen 'satte vogna framfor hesten." (5) A. Boyden påpekte i 1947 at ND homologi først krevde at vi fant opphavet og så besluttet at de korresponderende organer eller deler" er homologe. "Som om vi kunne vite opphavet uten de essensielle likhetene til å lede oss!" (6) Da ND-paleontolog G.G. Simpson forsøkte å benytte homologi som begrunnelse for felles stamfar, beskyldte biologer R. Sokal og P. Sneath ham for 'sirkulært resonnement.' (7)
Til tross for dette forsøker mange tilhengere av ND å forsvare sin bruk av homologi mot anklagen om sirkel-resonnement. I 1966 påpekte M. Ghiselin at ND-definisjoner ikke er sirkulære fordi homologi ikke er definert ut fra seg selv. (8) Men selv om begreps-definisjonen ikke er det, så bygger underliggende resonnement på et sirkulært resonnement. Året etter argumenterte D. Hull at begrunnelsen ikke er sirkulær, men bare et eks. på den vitenskapelige metoden med 'suksessiv approksimering' (9) i følge Hull starter evolusjonære biologer med å anta en bestemt avstamnings-hypotese, og så benyttes likheter til å raffinere denne hypotesen. Men metoden, som av datidens kritikere ble kalt 'groping' funger kun, om i det hele, ved å anta at felles avstamning er tilfelle. Dersom spørsmålet er hvorvidt ND holder mål, så er Hulls approksimering bare et annet sirkulært argument.
Kontroversen har rast siden. ND fortsetter å forsvare sin forbindelse fra homologi til felles avstamning, mens kritikere hevder at det forvirrer definisjonen med forklaring, og leder til sirkulært resonnement. R. Brady en uttalt kritiker av ND i siste generasjon, observerte: "Ved å gjøre vår forklaring til definisjon av av tilstanden som skal forklares, uttrykker vi ikke en vitenskapelig hypotese, men tro. Vi er så overbevist at vår forklaring er sann, at vi ikke lenger ser behov for å skjelne den fra situasjonen vi forsøker å forklare. Dogmatiske forsøk av dette slaget, må til slutt forlate den vitenskapelige sfære." (11)
Bilde 7. Forutsetter det som skal vises
Referanser:
3. Ch. Darwin 'On the Origin of Species', 6.utg. (London: John Murray, 1972); 403
4. E. Mayr, The Growth of biological Thougth (Cambridge, Mass: Harvard Univ. Press, 1932), 232,465
5. J. H. Woodger, "On Biological Transformations," In W.E. Le Gros Clark og P.B. Medawar (forf.) Oxford: Clarendon Press, 1945) 109
6. A. Boyden, "Homology and Analogy, " American Midland Naturalist 37 (1947): 648-669
7. R.R. Sokal og P. H. S. Sneath, Principles of Numerical Taxonomy (San Fr: Freeman, 1963), 21
8. M.T. Ghiselin, "An Application of the Theory of Definitions to Systematical Principles," Systematic Zoology 15 (1966): 127-130
9. David L. Hull, "Certainty and Circularity in Evolutionary Taxonomy," Evolution 21 (1967): 174-189
11. R. H. Brady, "On the Independence of Systematics," Cladistics 1 (1985): 113-126
Oversettelse og bilder ved Asbjørn E. Lund
{kind=link}